
a
b
c
d
e
f
g
h
i
j
k
l
m
n
o
p
q
r
s
t
u
v
w
x
y
z
#
Hematology

Adverse Effects of Transfusion
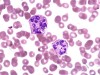
Arneth Count for Neutrophils: Principle, Procedure, and Clinical Significance

Bleeding Time (BT) and Clotting Time (CT)

Blood Clotting Process
Blood Group Systems

Blood Screening Market

Causes of Erroneous Results (Interferences Causing Abnormal Result) With Hematology Analyzer

Common Applications of Flow Cytometry in Hematology
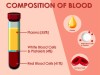
Composition of Blood and Its Functions

D-Dimer Test and Its Clinical Significance

Determination of Blood Group by Slide Method

Determination of Blood Group by Tube Method

Determination of WBC Differential Count by Microscopic Method
