Ancient Choanoflagellate Gene Reprograms Mouse Cells, Producing Mosaic Mice
Ancient single‑celled organism genes reprogram mouse cells, shaping embryonic development and showing a billion‑year evolutionary link.
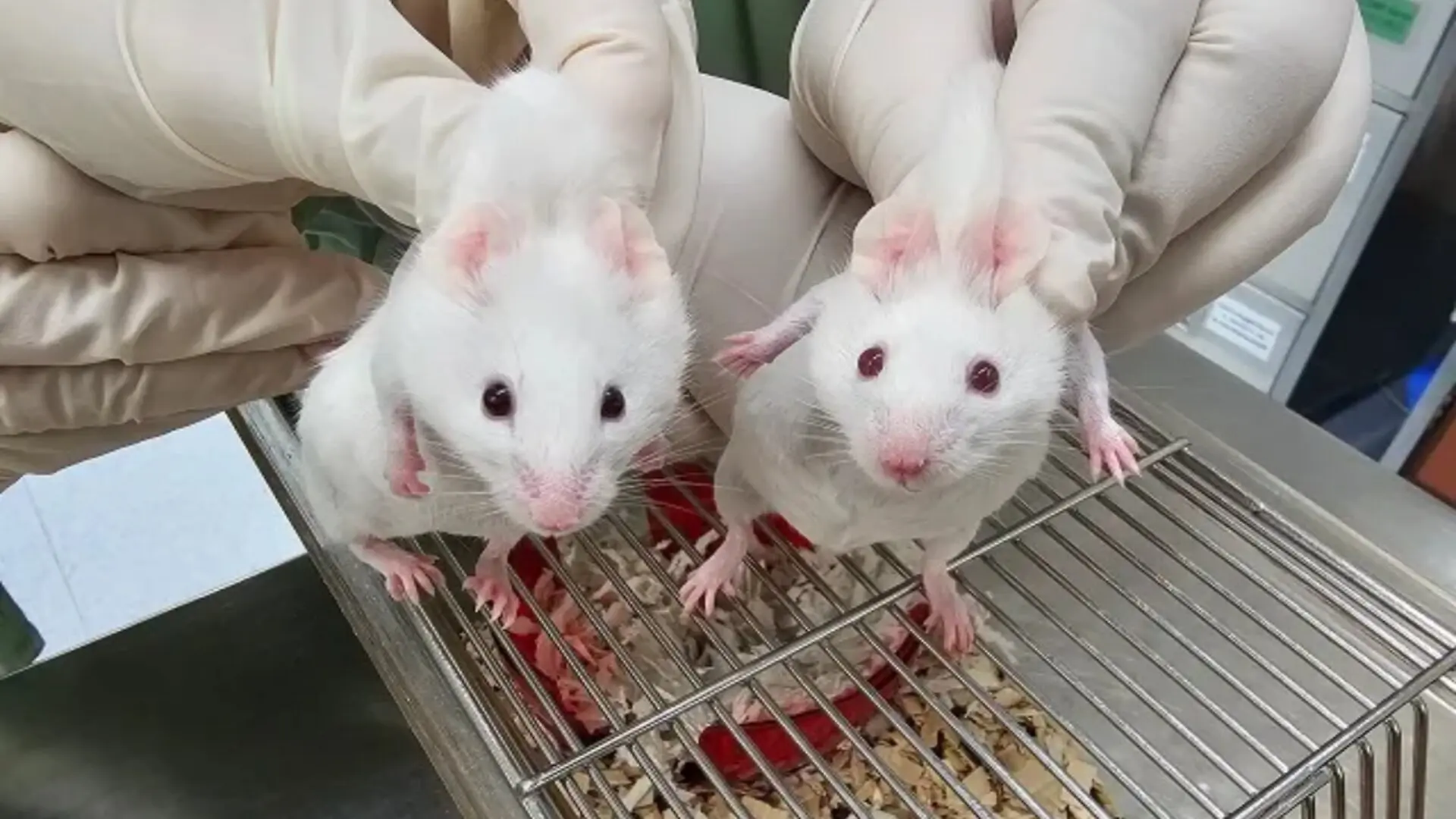
Researchers have tapped a genetic mechanism that predates animal life to rewire mouse cells, steering embryonic development and yielding chimeric mammals that display distinct inherited characteristics, as reported in Nature Communications.
A Reverse Evolution Experiment
The investigation flips conventional developmental biology on its head: ancient genes, typically found in single‑celled organisms, were inserted into mouse cells and sparked a cascade of changes normally reserved for complex multicellular organisms. The team focused on choanoflagellates—microscopic eukaryotes regarded as the closest living relatives of animals—and examined their Sox‑like regulatory genes, which were once thought to be exclusive to multicellular life. When these primordial genes entered mouse cells, they did more than persist as relics; they actively remodeled cellular identity, driving cells into a pluripotent state capable of forming diverse tissue types.
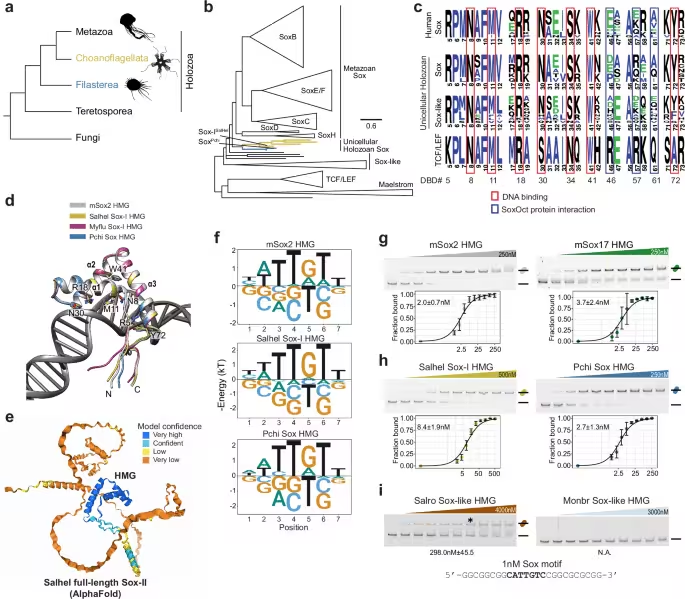
These engineered cells were coaxed into induced pluripotent stem cells that later merged with developing embryos. The resulting mice exhibited visible mosaics—black fur patches and dark eyes—signifying the integration of genetically distinct cell lines. The authors interpret the outcome as evidence that core mechanisms governing cellular flexibility have been conserved for nearly a billion years, challenging prevailing ideas about the timing of developmental gene emergence.
Re‑engineering Stem‑Cell Pathways with Ancient Genes
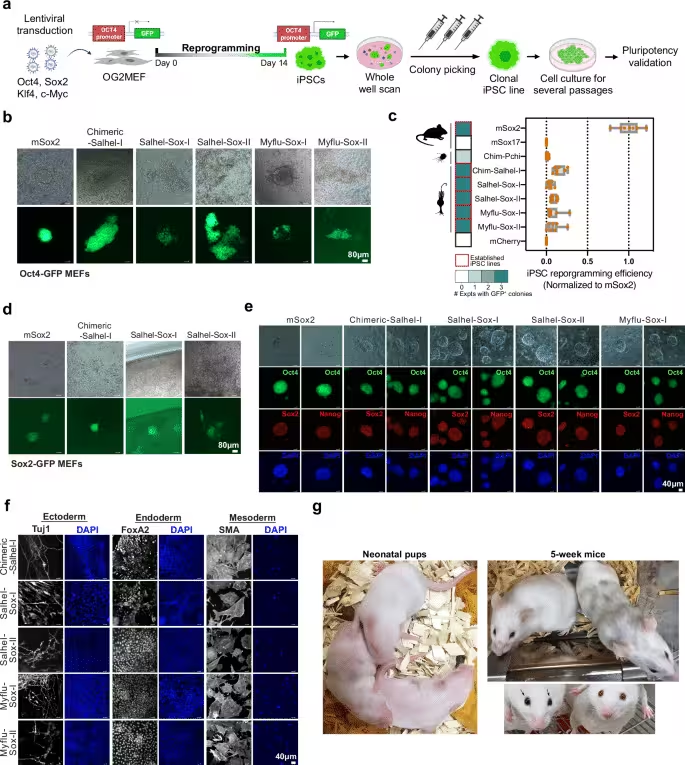
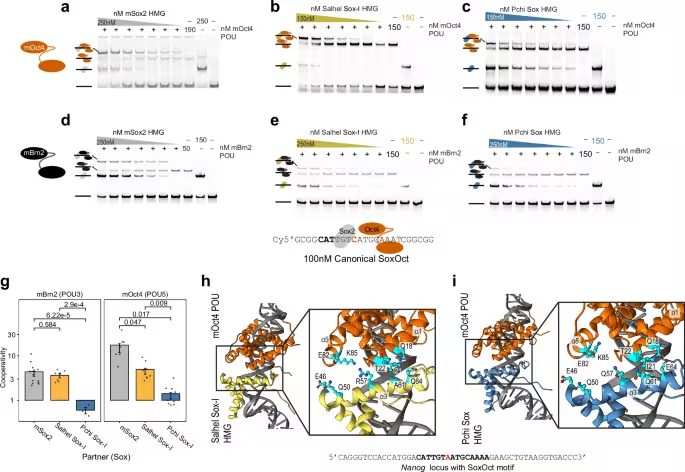
In the reported study (Nature Communications), scientists swapped a pivotal mammalian stem‑cell regulator for a choanoflagellate counterpart. Although choanoflagellates lack true stem cells, they possess Sox‑like genes previously believed to be animal‑specific. When these genes entered mouse fibroblasts, they ignited pluripotency circuits that normally rely on mammalian transcription factors. The induced pluripotent stem cells were subsequently introduced into early embryos, where they contributed to multiple lineages.
Mosaic animals emerged, confirming that the ancient genes exerted functional influence throughout development. The authors highlight the surprising compatibility between distant evolutionary lineages, suggesting that modern developmental processes still lean on deeply conserved molecular scaffolds.
Reflecting on the work, study author Dr Alex de Mendoza noted,
“By successfully creating a mouse using molecular tools derived from our single‑celled relatives, we’re witnessing an extraordinary continuity of function across nearly a billion years of evolution.”
The authors also propose that genes linked to stem‑cell behavior may have arisen before stem cells themselves, prompting a rethink of early genetic regulation.
Implications for the Origin of Cellular Plasticity
Beyond the technical breakthrough, the study raises fundamental questions about the emergence of biological complexity. Choanoflagellates lack true stem cells, yet they harbor genetic elements that mirror those used in animal development. This paradox hints that molecular systems associated with multicellular organization may have originated in a unicellular context and later been repurposed as organisms grew more complex. As Dr de Mendoza explained,
“Choanoflagellates don’t have stem cells, they’re single‑celled organisms, but they have these genes, likely to control basic cellular processes that multicellular animals probably later repurposed for building complex bodies,”
The presence of such genes in single‑celled life supports the idea that evolutionary innovation often stems from the reuse of existing components rather than from de novo invention. Cellular identity, differentiation, and developmental control may therefore rest on ancient regulatory frameworks that predate animals by hundreds of millions of years. This perspective shifts focus from abrupt evolutionary leaps toward gradual adaptation of long‑standing molecular architectures, blurring the genetic boundary between unicellular and multicellular organisms.
Chimera Formation and Developmental Flexibility
One of the most striking visual outcomes was the generation of chimeric mice—organisms composed of cells from distinct genetic origins. After injecting the engineered stem cells into embryos, they integrated across multiple tissues, producing observable mosaics in coat and eye coloration that served as biological markers of cellular contribution. The stability of these reprogrammed cells within a living organism demonstrates that pluripotent states can be sustained beyond the early embryo.
The authors note that such compatibility underscores the robustness of developmental systems, which can accommodate genetically diverse inputs without compromising overall growth. Dr Ralf Jauch added,
“Studying the ancient roots of these genetic tools lets us innovate with a clearer view of how pluripotency mechanisms can be tweaked or optimised,”
Overall, the work suggests that developmental plasticity is a deeply rooted biological trait, extending far beyond the origins of animals and offering new avenues for biomedical research.
This article has been fact checked for accuracy, with information verified against reputable sources. Learn more about us and our editorial process.
Last reviewed on .
Article history
- Latest version
Reference(s)
- Gao, Ya. “The emergence of Sox and POU transcription factors predates the origins of animal stem cells - Nature Communications.”, vol. 15, no. 1, November 14, 2024, pp. 9868 Nature, doi: 10.1038/s41467-024-54152-x. <https://www.nature.com/articles/s41467-024-54152-x>.
Cite this page:
- Posted by Rohan Kumar