Arc Protein Hijacked to Spread Toxic Tau: New Pathway Offers Fresh Alzheimer’s Target
Study finds a brain protein may help Alzheimer’s spread via tiny vesicles, offering a fresh target to slow disease progression.
New research published in Cell points to a previously unknown cellular route that may change how scientists view the spread of Alzheimer’s disease across the brain. The study identifies a normal brain protein that appears to unintentionally ferry toxic Tau, the protein closely linked to neuronal loss and memory decline, between cells. The findings imply that disease progression depends not only on the accumulation of harmful proteins inside neurons but also on the mechanisms that move these proteins from one cell to another.
By facilitating intercellular transfer, this pathway could speed the propagation of pathology throughout different brain regions over time. Experiments in laboratory models showed that the protein Arc plays a pivotal role in packaging and shuttling Tau from one neuron to the next, suggesting that a communication system essential for healthy brain function may be co‑opted in disease.
Arc Bridges Normal Signalling and Disease Transmission
The investigation, detailed in Cell, highlights Arc as a double‑edged molecule that supports both regular neuronal activity and the spread of Alzheimer‑related damage. Under normal circumstances, Arc helps neurons exchange signals via tiny membrane‑bound packets called extracellular vesicles, which travel across synapses to maintain brain homeostasis.
In Alzheimer’s models, the researchers found that toxic Tau hijacks this vesicle system. Rather than staying confined to the originating cell, Tau binds to Arc‑laden vesicles and is delivered to neighboring healthy neurons, where it can trigger further protein misfolding and cellular impairment. The data suggest that a network designed for communication can become a conduit for disease spread.
Comparisons between normal mice and those lacking Arc revealed a dramatic drop in Tau movement when Arc was absent, underscoring its central role in intercellular transmission.
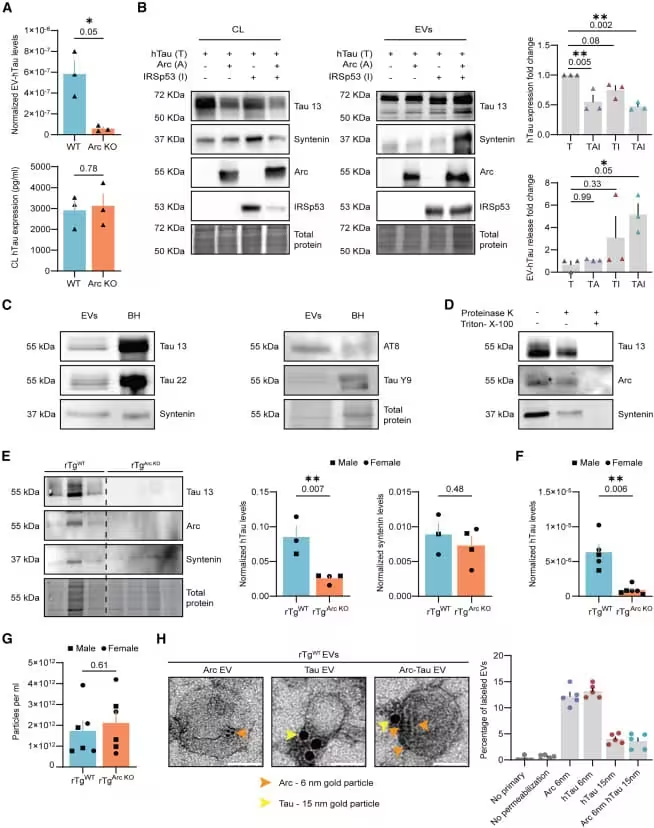
How Toxic Tau Forms “Glue‑Like” Aggregates Inside Neurons
Once initiated, the process can cascade through neural networks, spreading damage in a ripple effect. The study demonstrates that Arc‑associated vesicles may act as carriers for these seeding fragments. Inside the vesicles, Tau is shielded from enzymatic breakdown, increasing the likelihood that it reaches new neurons. This mechanism offers a cellular explanation for the progressive loss of memory and cognition observed in Alzheimer’s patients.
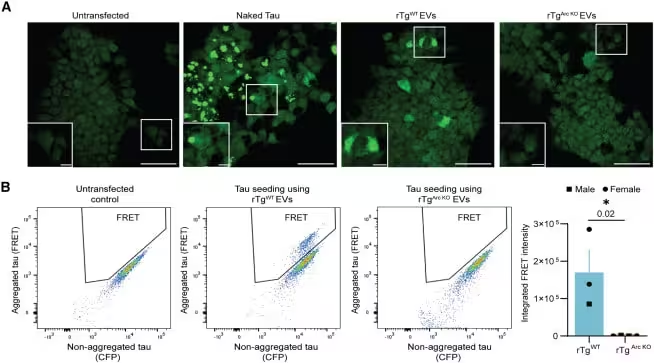
Mouse Experiments Show Sharp Decline in Tau Spread Without Arc
Data from mouse models support the proposed mechanism. Researchers compared brains that express normal levels of Arc with those where the protein was genetically removed. In the Arc‑deficient mice, the transfer of Tau between neurons fell dramatically, altering the trajectory of disease development. One of the investigators summed up the observation: “When we removed Arc, we saw that the transfer of Tau was severely, severely reduced. It was almost gone.” These results indicate that Arc may be required for efficient intercellular movement of Tau.
In the absence of Arc, toxic proteins tend to remain trapped inside affected neurons, potentially leading to higher intracellular toxicity while limiting spread to neighboring cells. This trade‑off highlights the challenge of designing therapies that block disease propagation without harming the neurons that rely on Arc for normal waste management.
Balancing Neuronal Protection and Pathology Propagation
Further analysis revealed a nuanced role for Arc in neuronal health. When active, Arc enables neurons to export excess Tau, reducing internal stress and extending cell survival. At the same time, the exported material can be taken up by nearby healthy cells, extending the pathological cascade.
These findings suggest that directly inhibiting Arc could worsen damage in already compromised cells. Consequently, researchers are exploring strategies that intercept vesicles after they leave the donor neuron rather than preventing their formation altogether.
Potential Impact on Future Alzheimer’s Therapies
The team also detected Arc‑ and Tau‑laden vesicles in human brain samples, hinting that the mechanism may operate beyond animal models. Nevertheless, the authors caution that many questions remain about how this process functions in people. “Most of the work we’ve been doing is in mice, not in humans,” Shepherd notes. “We have some clues that whatever is happening in these mice could also be happening in humans, but we don’t know that yet. And we’re far away from saying that we’re developing a treatment for anything.”
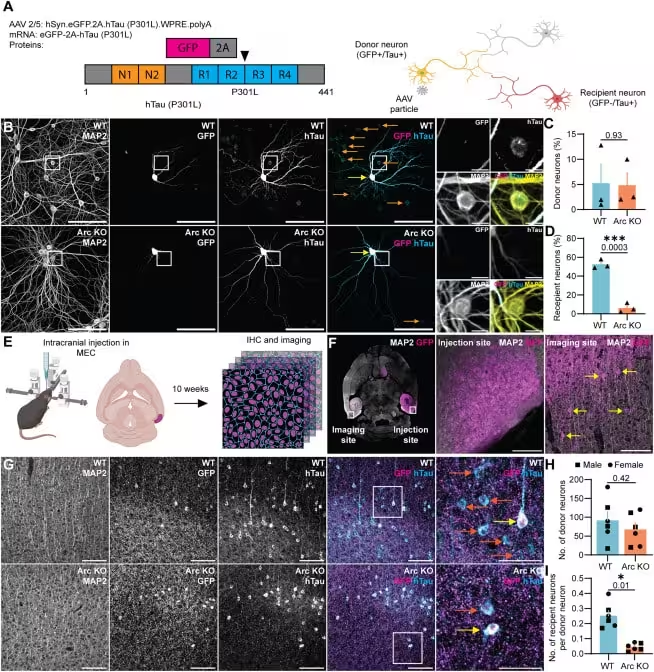
Shepherd adds that targeting the vesicles after they exit the donor cell could provide a therapeutic window: “If we could target these particular EVs, that would be a really useful therapy strategy. For someone with early‑onset Alzheimer’s or dementia, if we could stop the spread, then we could prevent further damage and cognitive decline.” This approach shifts the focus from removing toxic proteins to blocking their movement, opening a new avenue in Alzheimer’s research.
This article has been fact checked for accuracy, with information verified against reputable sources. Learn more about us and our editorial process.
Last reviewed on .
Article history
- Latest version
Cite this page:
- Posted by David Anderson